分享至
分享至



单细胞分析的丝滑IMAX体验: icpTOF 以多元素指纹量化海藻细胞与纳米颗粒间相互作用为例
ICP-TOF
电感耦合等离子体-飞行时间质谱

L Hendriks et al.
确定细胞中金属元素的生物利用率的传统方法一般需对细胞进行酸消解,然后利用溶液进样电感耦合等离子体质谱(ICP-MS)进行后续分析。这种方法的缺点是需要大量的细胞,并且只能为给定的细胞群体提供平均值1。众所周知,千人千面,不同群体以及同群体细胞的特异性在文献中也多有报道2。基于这个大前提,使用特定的分析方法对不同群或同群细胞进行逐序单个分析,获取与单个细胞特异性有关的大数据就尤其重要(见图1)。本文中介绍的单细胞-电感耦合等离子体质谱法(sc-ICP-MS)与之前介绍过的单颗粒ICP-MS(sp-ICP-MS)基本类似(微信公共号:粒粒皆信息:什么是单颗粒物ICP-MS质谱分析法?)。事实上,上述两种技术都依赖于相同的基本原理和icpTOF瞬时事件全谱多元素测量能力,从而可以获得由单一个体产生的微秒时间区间内的瞬时信号,例如单个纳米颗粒(NPs)或单个细胞。这等同在拍一段有很多快速武术对打的电影场景,需要使用高速摄像机来捕捉每一个武打动作细节和变化,同时也不漏过颜色,声音等关键信息,这样才能最终呈现出高清120Hz的作品。
单颗粒ICP-MS方法的基础概念和硬件构架3源于2003年Degueldre等发表的第一篇论文。在过去的二十年间,通过进样系统,数据采集硬件和数据处理专用软件的进一步发展和商业化,不断增加的科研文献见证了该技术领域的迅速成熟。在单颗粒ICP-MS上投入的研究和应用开发同样的也使单细胞ICP-MS分析受益。
在单细胞ICP-MS中,细胞悬浮液经超声波雾化后形成的液滴被带入ICP-MS等离子体中。细胞在等离子体中依次被汽化、原子化和最终离子化。每个细胞产生一个含有多种元素的离子云,在仪器上被检测为高于背景的时长几百微秒的单个信号峰。与单颗粒ICP-MS类似,记录到的尖峰频率与细胞数量浓度成正比,这些尖峰的强度则与细胞中该元素质量有关。这种技术已经成功的应用在测定海藻中的镁元素含量4,并进一步用于纳米颗粒物毒理学研究中评估细胞对纳米颗粒物的摄取情况5,6,7。
虽然单细胞ICP-MS的测量方法看起来很简单,但要获得真实可靠的数据,实施起来需要注重的细节很多。除了需要额外注意来自培养基的可能高背景信号和细胞在样品导入系统中的潜在破损,在单细胞研究中反复报道的一个主要瓶颈是细胞进样装置的低运输效率,这是因为与纳米颗粒物相比,细胞的尺寸更大,在传输过程中也更容易损失。事实上,传统的系统通常包括一个旋风式雾化室,是专为引入较小的溶液液滴而设计的,导致细胞传输效率低于10%。而用于单细胞导入的定制系统,包括改进的雾化器或全消耗喷雾室8,9,以及其他创新设计10,11,经过多年反复测试,已被验证可以高效传输单细胞进入ICP-MS。
另一个瓶颈在于质谱仪器质量分析器的性能:传统的ICP-MS仪器具有单四极杆或扇形场质量分析器,在进行单细胞分析时最多只能同时检测一到两种元素信息(只能拍黑白影片)。而在常见的单颗粒分析场景中,比如在纳米毒理学研究中,在试图量化纳米颗粒物(特征金属元素)和细胞(蛋白固有元素)的关联时,需要同时获得单细胞事件内多种元素浓度信息。为了获得微秒级事件信息全貌,快速且广谱分析的质量分析器,如飞行时间质量分析器等高精尖‘摄影器材’是必不可少的(提供高清彩色120Hz影片给观众更加真实的IMAX观影体验)。

图1:a)在对细胞进行酸消解后,通过传统的雾化法将溶液样品引入ICP-MS,并记录仪器获得的稳态信号。这种整体分析法对初始样品中所包含的数千个细胞获得一个平均值。然而这种实验是基于细胞是均匀的假设,而忽略了细胞具有多样性的事实。因此,少数细胞群(用绿色和紫色表示),在元素组成上虽与主类细胞有差异,却没有被体现在结果中,这完美的诠释了辛普森悖论。b)在单细胞ICP-MS方法中,将细胞悬浮液稀释后,在单位时间内仅有一个细胞个体被引入ICP-MS等离子体。每个细胞产生一个独立的离子云,作为信号峰被ICP-MS仪器记录。这种方法允许检测每一个单独的细胞,从而保证了细胞特异性信息的无损获取和保存。简单来说,在单细胞ICP-MS中,细胞是以个为单位进行分析的,可以根据它们不同的分析物含量识别出不同的群体,而不是仅仅产生一个平均值。
icpTOF
飞行时间质谱法
在飞行时间质谱法(TOF-MS)中,其基本原理是根据离子到达检测器前通过固定长度的飞行管的飞行时间来精确分辨离子。离子束在脉冲加速电压后具有相同的动能,但轻的离子会比重的离子获得更高的速率,进而更早到达检测器。测量所有离子的陆续到达时间可以得到一个连续时间谱,经过简单的校准和换算后可以得到一张全质谱谱图(一般6-280 Th)。TOF质量分析仪的主要优点是:对分析的元素及同位素的数量没有限制,而且全谱数据采集速度快(通常几十微秒就可以获得一张全元素谱图)。这样的快速全谱数据采集能力在处理单一实体(如单细胞)检测时尤其重要,因为单细胞产生的瞬时事件长度很短,一般在200-500微秒区间。
飞行时间技术在单细胞分析领域并不是一个新概念,最初是由Bandura在2009年提出的,其原型机12用于单个细胞的时间分辨分析13,从而为众所周知的 "质谱流式 "领域打开了大门。这项应用使用稳定的稀土金属同位素来标记细胞,从而允许通过其金属标记物来检测相应细胞14。除了展现了生物研究和药物筛选应用中的巨大潜力,质谱流式也被用于检测细菌细胞中的银纳米颗粒15。然而,由于质量检测范围有限(>80 Da)和涉及染色的样品制备程序,质谱流式细胞技术无法检测许多固有元素。
与质谱流式不同的,如图2a) 所示的ICP-TOF (TOFWERK AG, 瑞士) 可以测量从质荷比6到280的全谱图16,从而可以覆盖轻质元素,如Na, Mg, P, S, K, Ca, Mn, Fe, Cu, Zn等。这些元素是活细胞的固有元素,它们的分布(也被称为细胞离子组17)可以作为细胞发育状态的指标18。例如,磷存在于核酸(DNA和RNA)中,也是ATP、CTP、GTP和UTP等能量化合物的重要成分。钠和钾在电信号的传输中起作用,而锌被不同的生物过程中的多种酶用作催化剂。由于ICP-TOF-MS的同时多元素检测能力,可以在多种元素的相关分析基础上进行指纹识别19。如图2b) 所示,镁、磷、锰、铁、铜和锌被鉴定为被分析藻类的本征指纹元素。不需要标记或染色,即可依据细胞的 "天然 "元素指纹来进行单细胞分析20,21。通过测量特定细胞类型的金属微量元素,则可以获得更细致的指纹信息。例如,海藻细胞富含镁等金属微量元素,镁是叶绿素的核心组成部分,对光合作用至关重要。因此,金属微量元素的组成可以作为一种独特的指纹来明确识别不同的细胞种类。通过测量单细胞的金属元素组分,可更好地了解由金属蛋白和金属酶调节的基本生物过程,从而解密细胞生命周期不同状态22。尽管细胞的生物化学并不完全反映在其离子组上,但通过监测其金属含量的变化,可以确定地获得对细胞状况和生物过程的更深入了解。
通过使用TOF质量分析仪作为检测器,可以动态系统地获得完整的质谱数据,从而可以对发现特定实体本身及其所处环境进行连续或高通量表征。因此在纳米毒理学背景下,人们可以很容易地确定纳米颗粒物是否与细胞相关联。
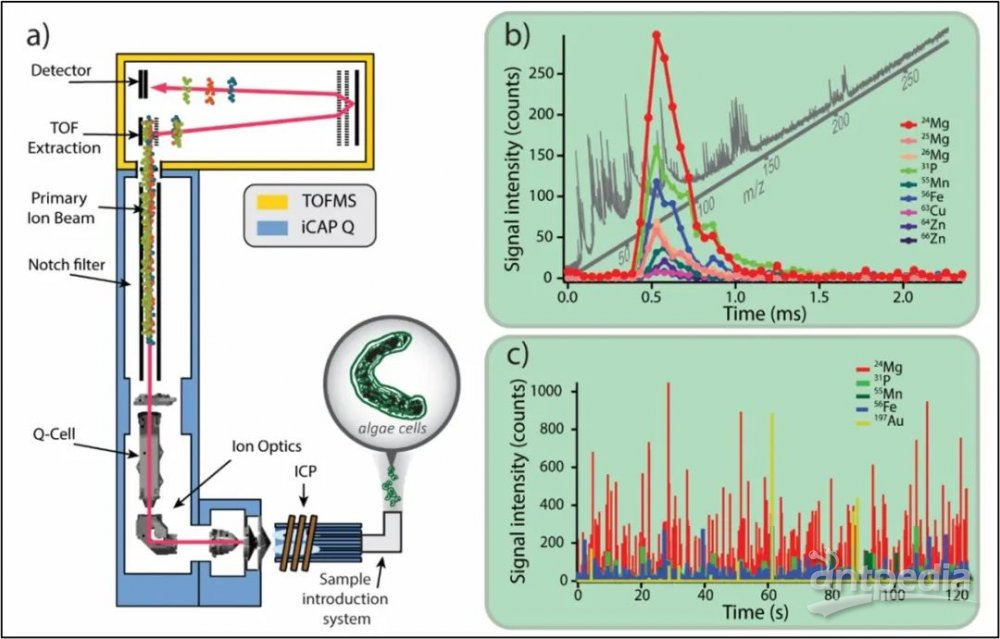
图2:a) icpTOF仪器(TOFWERK AG, Thun, Switzerland)的示意图:在iCAP Q(Thermo Scientific, Bremen, Germany)的框架上搭配一套高分辨率飞行时间质量分析器。因此,ICP-TOF受益于与iCAP Q相同的ICP离子源、离子光学、碰撞/反应池技术和样品引入设备。b) 用48 µs时间分辩率采集的淡水藻类细胞raphidocelis subcapitata的瞬时信号速率。c) 藻类细胞通常用于毒理学风险评估研究,这里在暴露于金纳米颗粒一段时间后进行分析,以调查其摄取情况。在ICP-TOF的全质量数范围内,可以根据检测细胞的本征元素指纹对细胞进行追踪,并能直接定量测量纳米颗粒物-细胞的关联。

icpTOF
单细胞分析应用实例
单一实体分析,与批量样品测量相比,能产生信号的质量相对有限,这对仪器灵敏度要求更高。下面的应用案例研究展示了icpTOF S2仪器(TOFWERK AG,瑞士)的性能指标:具有与单四极杆ICP-MS类似的高灵敏度,又可同时快速检测全谱信号,特别适合分析单一实体,如单细胞或纳米颗粒(NPs)等。随着工业和日常生活中纳米颗粒物的广泛使用,纳米安全和纳米毒理学在过去20年一直是深入研究的课题。纳米颗粒物的安全评估研究中的一个重要参数是其在细胞摄取的分析和量化。
透射电子显微镜(TEM)和扫描电子显微镜(SEM)具有高空间分辨率,它们经常被用于细胞内纳米颗粒物的分析23,24。尽管有令人印象深刻的成像能力,基于电子显微镜方法的一个主要缺点是对样品制备的繁琐要求。此外,由于没有额外的元素定量或自动图像分析,获得的图像是定性的且结果较难被解读25,26。如前所述,单细胞ICP-MS也可用于量化细胞对纳米颗粒物的摄取,根据观察到的信号峰的强度大小,提供与细胞相‘关联’的纳米颗粒数量的信息5,6。这类实验通常有以下三个明显的观察结果:
· 只检测到纳米颗粒物中的特征元素,表明溶液中存在纳米颗粒物
· 只检测到细胞固有元素而没有任何纳米颗粒物中的元素,表明细胞并没有与纳米颗粒物相关联
· 同时检测到细胞固有元素和纳米颗粒物中的元素,意味着两者有关联
根据观察到的相关联的纳米颗粒/细胞峰的频率和幅度,可以确定摄取了纳米颗粒物的细胞的百分比以及与每个藻类细胞相关的纳米颗粒数量的估计值。在理想的情况下,可以根据浓度和暴露时间动态地对海藻细胞和纳米颗粒数量的相关性的进行评估。
在本案例研究中,将海藻细胞暴露在BaSO4(NM-220)溶液中72小时,接着按照Merrifield等人提出的程序进行清洗5,去除未与细胞结合的纳米颗粒。在暴露后并在ISO8692藻类培养基中进行冲洗后27,样品中预计只包含与藻类细胞相关联的纳米颗粒物。随后,样品被储存在15毫升的试剂管中,用锡纸包裹,等待分析。
在使用四极杆ICP-MS(NexION 350D,PerkinElmer,Shelton,CT,USA)进行单细胞的初始研究中,我们发现清洗后的细胞悬浮液中仍存在BaSO4纳米颗粒,如图3a所示。有学者认为未关联的纳米颗粒已经去除,而这些检测到的纳米颗粒是与海藻细胞相关联的。然而由于只测量了一种元素138Ba,并不能完全证实这一猜想。
我们使用单细胞ICP-TOF-MS(见图2a)重复了一个类似的实验。从图2b中我们可以知道被分析的藻类细胞的本征元素指纹,即只有同时检测到Mg、P、Mn和Fe等元素时才被认为检测到了藻类细胞。令人惊讶的是,即使暴露72小时后,BaSO4 纳米颗粒与水藻细胞的指纹信号没有显著关联(图3b)。可以看到,Ba仅与Mg和Fe的信号同时被检测到,而没有水藻的其他指纹信号同时出现。虽然缺失的元素信号强度有可能是低于仪器检测极限,但至少这说明检测到的元素与藻类细胞的本征元素指纹不一致。然而在检测到藻类细胞的指纹信号中,没有观测到Ba元素信号。综上所述,如果没有icpTOF瞬时多元素检测能力,在清洗后细胞悬浮液中检测到的纳米颗粒的Ba信号很容易被误解为是与藻类细胞相关联的颗粒物。

图3:a)实验流程图。在样品暴露于纳米颗粒物72小时后,细胞被清洗以去除上清液中游离态的纳米颗粒物。b) 通过使用飞行时间质谱仪重复单细胞测量,可以跟踪细胞的元素指纹,以验证纳米颗粒物信号和细胞信号的是否同时出现。结果显示虽然纳米颗粒物和细胞没有直接关联,但Ba信号与Mg和Fe信号是一起出现的。
这些结果导致了对可能引发该现象的机制的讨论。一个合理的解释是海藻细胞通过释放胞外聚合物物质(EPS)来清除粘附在细胞表面的纳米颗粒物。EPS被认为是影响藻类细胞对纳米颗粒的生物利用率的关键因素28,29。EPS产量的增加可使藻类细胞主动脱落纳米颗粒,从而减轻摄取或吸附到细胞外部,而纳米颗粒仍然以被包含在EPS中的形式存在于溶液中。虽然缺乏关于这种行为的定量数据,但足以解释BaSO4纳米颗粒信号与Mg和Fe信号的契合。当然Fe与Ba信号的同时出现还可以被解释为溶解的Ba与ISO 8692培养基中的EDTA络合在了一起,而EDTA被添加在溶液中以保持Fe的生物可利用率。要回答这个问题,我们使用TEM观察到EPS聚集体中包裹有纳米颗粒(图4)。由于TEM局限于定性分析,再加上EPS结构微妙,这种包裹的确切机制和发生频率很难被量化。然而单细胞ICP-TOF-MS则可以直接对这一现象进行定量分析,而不需要对样品进行复杂的制备,同时还可以在较短的时间内分析更多的藻类细胞及EPS聚集体,提供更可靠的统计数据。此外,单细胞ICP-TOF-MS可以动态地从藻类悬浮液中不间断取样,评估这种清除行为的发生频率与样品浓度和时间的关系,进一步了解藻类细胞和纳米颗粒之间的相互作用。这种利用ICP-TOF研究动态摄取和清除行为的研究思路不仅限于藻类细胞,还可以扩展到纳米医学或纳米生物技术的其他类型细胞,如哺乳动物细胞或细菌。

图4:一个藻类细胞(Raphidocelis subcapitata)的透射电子显微镜图像,该细胞之前暴露在银纳米颗粒物中,脱落的细胞外聚合物物质(EPS)含有银纳米颗粒。(由Louise H. S. Jensen和Sara N. Sørensen提供)。
正如本研究强调的那样,尽管传统的四极杆质谱(sc-ICP-MS)可以测量单细胞,但它最多只能同时测量一种或两种元素或同位素,所以即使检测到纳米颗粒信号也不能100%确定其与细胞直接关联。另外还需要TEM来确定颗粒物是否被藻类吸收在内部或简单附着在细胞外部。然而使用ICP-TOF-MS可以将被暴露在纳米颗粒物中藻类的离子组与对照藻类的离子组进行比较,从而评估它们的状况。这些信息对于从机理上理解海藻细胞与纳米颗粒物的相互作用非常有价值,并可以进一步促进开发以生理学为基础的纳米颗粒物风险评估工具。
icpTOF
结论与展望
单细胞ICP-TOF-MS是一个新兴的、令人兴奋且快速发展的研究领域。虽然尚需数年时间才能达到质谱流式技术在单细胞多参数分析方面的水平,但ICP-TOF-MS得益于灵敏度的提高和同时全质谱检测能力,能够基于元素指纹检测未被标记的细胞,从而为新的实验设计创意提供可能性。例如,除了测量纳米颗粒物和细胞的相关性外,ICP-TOF-MS记录的多元素数据可用于评估细胞在纳米颗粒介导毒性影响下的不同状态。
除了液体样品引入方法之外,也可以使用激光剥蚀(LA)-ICP-TOF-MS进行单细胞分析30,31。通过将细胞放在样品台上并使用激光扫描,可以产生整个细胞层面上的元素分布二维图像,其中每个像素包含一个完整的全谱信息。LA-ICP-TOF-MS成像的高空间分辨率对纳米毒理学研究特别有意义,因为它可以观察和定位纳米颗粒物在亚细胞结构中的聚集,以进一步了解各种现象(如摄取、积累和释放纳米颗粒)。
此外,所生成的大量数据可以通过降维技术进行处理,如主成分分析(PCA)或机器学习工具,并提取与细胞状态和类型有关的信息,从而使细胞的分类变得更容易。这在质谱流式工作流程中是常见的处理方法。这项技术不仅限于纳米毒理学研究,还可以扩展到金属组学和细胞生物学中。无论如何,我们将继续努力改进飞行时间质谱ICP-TOF-MS技术,使其在更广阔的应用领域发挥作用。
icpTOF
致谢
作者感谢Olga Meili和Aiga Mackevica校对本文并提供反馈。Lars M. Skjolding得到了PATROLS – Advanced Tools for NanoSafety Testing项目资助(760813)。感谢Louise Helene Søgaard Jensen和Sara Nørgaard Sørensen允许使用图4中的TEM图像。最后特别感谢Robert Thomas邀请在Spectroscopy杂志中的 "原子视角专栏 "刊登此文。
原文链接:https://www.tofwerk.com/wp-content/uploads/Atomic-Perspectives-October.pdf (请点击左下角“阅读原文”跳转)
参考文献
1 S. J. Altschuler and L. F. Wu, Cell, 2010, 141, 559–563.
2 W. M. Elsasser, Proc. Natl. Acad. Sci. U. S. A., 1984, 81, 5126–5129.
3 C. Degueldre and P. Y. Favarger, Colloids Surfaces A Physicochem. Eng. Asp., 2003, 217, 137–142.
4 K. S. Ho and W. T. Chan, J. Anal. At. Spectrom., 2010, 25, 1114–1122.
5 R. C. Merrifield, C. Stephan and J. R. Lead, Environ. Sci. Technol., 2018, 52, 2271–2277.
6 F. Abdolahpur Monikh, B. Fryer, D. Arenas-Lago, M. G. Vijver, G. Krishna Darbha, E. Valsami-Jones and W. J. G. M. Peijnenburg, Environ. Sci. Technol. Lett., 2019, 6, 732–738.
7 I. L. Hsiao, F. S. Bierkandt, P. Reichardt, A. Luch, Y. J. Huang, N. Jakubowski, J. Tentschert and A. Haase, J. Nanobiotechnology, 2016, 14, 1–13.
8 A. S. Groombridge, S. I. Miyashita, S. I. Fujii, K. Nagasawa, T. Okahashi, M. Ohata, T. Umemura, A. Takatsu, K. Inagaki and K. Chiba, Anal. Sci., 2013, 29, 597–603.
9 M. Corte-Rodríguez, R. Álvarez-Fernández García, P. García-Cancela, M. Montes-Bayón, J. Bettmer and D. . Kutscher, Curr. Trends Mass Spectrom., 2020, 18, 6–10.
10 K. Shigeta, H. Traub, U. Panne, A. Okino, L. Rottmann and N. Jakubowski, J. Anal. At. Spectrom., 2013, 28, 646–656.
11 P. E. Verboket, O. Borovinskaya, N. Meyer, D. Günther and P. S. Dittrich, Anal. Chem., 2014, 86, 6012–6018.
12 D. R. Bandura, V. I. Baranov, O. I. Ornatsky, A. Antonov, R. Kinach, X. Lou, S. Pavlov, S. Vorobiev, J. E. Dick and S. D. Tanner, Anal. Chem., 2009, 81, 6813–6822.
13 K. R. Atkuri, J. C. Stevens and H. Neubert, Drug Metab. Dispos., 2015, 43, 227–233.
14 S. D. Tanner, V. I. Baranov, O. I. Ornatsky, D. R. Bandura and T. C. George, Cancer Immunol. Immunother., 2013.
15 Y. Guo, S. Baumgart, H. J. Stärk, H. Harms and S. Müller, Front. Microbiol., 2017, 8, 1–9.
16 L. Hendriks, A. Gundlach-Graham, B. Hattendorf and D. Günther, J. Anal. At. Spectrom., , DOI:10.1039/c6ja00400h.
17 M. Malinouski, N. M. Hasan, Y. Zhang, J. Seravalli, J. Lin, A. Avanesov, S. Lutsenko and V. N. Gladyshev, Nat. Commun., , DOI:10.1038/ncomms4301.
18 D. E. Salt, I. Baxter and B. Lahner, Annu. Rev. Plant Biol., 2008, 59, 709–733.
19 A. Praetorius, A. Gundlach-Graham, E. Goldberg, W. Fabienke, J. Navratilova, A. Gondikas, R. Kaegi, D. Günther, T. Hofmann and F. Von Der Kammer, Environ. Sci. Nano, 2017, 4, 307–314.
20 O. Borovinskaya, S. Aulakh and R. Markus, Tofw. appilcation note, 2019, 1–3.
21 M. von der Au, O. Borovinskaya, L. Flamigni, K. Kuhlmeier, C. Büchel and B. Meermann, Algal Res., 2020, 49, 101964.
22 L. Mueller, H. Traub, N. Jakubowski, D. Drescher, V. I. Baranov and J. Kneipp, Anal. Bioanal. Chem., 2014, 406, 6963–6977.
23 F. Piccapietra, C. G. Allue, L. Sigg and R. Behra, Environ. Sci. Technol., 2012, 46, 7390–7397.
24 F. Perreault, A. Oukarroum, S. P. Melegari, W. G. Matias and R. Popovic, Chemosphere, 2012, 87, 1388–1394.
25 L. H. S. Jensen, L. M. Skjolding, A. Thit, S. N. Sørensen, C. Købler, K. Mølhave and A. Baun, Environ. Toxicol. Chem., , DOI:10.1002/etc.3697.
26 C. Brandenberger, M. J. D. Clift, D. Vanhecke, C. Mühlfeld, V. Stone, P. Gehr and B. Rothen-Rutishauser, Part. Fibre Toxicol., , DOI:10.1186/1743-8977-7-15.
27 ISO, International Organization for Standarization. ISO 8692. Water quality - Fresh water algal growth inhibition test with unicellular green algae., 2012.
28 J. Zhao, X. Cao, X. Liu, Z. Wang, C. Zhang, J. C. White and B. Xing, Nanotoxicology, , DOI:10.1080/17435390.2016.1206149.
29 F. Chen, Z. Xiao, L. Yue, J. Wang, Y. Feng, X. Zhu, Z. Wang and B. Xing, Environ. Sci. Nano, 2019, 6, 1026–1042.
30 S. Theiner, A. Schoeberl, S. Neumayer and G. Koellensperger, J. Anal. At. Spectrom., 2019, 34, 1272–1278.
31 S. Theiner, A. Schweikert, C. Haberler, A. Peyrl and G. Koellensperger, Metallomics, , DOI:10.1039/d0mt00080a.
END